

How Can Viruses Change The Genetic Makeup
Full general Concepts
Genetic Modify in Viruses
Viruses are continuously changing equally a consequence of genetic selection. They undergo subtle genetic changes through mutation and major genetic changes through recombination. Mutation occurs when an error is incorporated in the viral genome. Recombination occurs when coinfecting viruses exchange genetic information, creating a novel virus.
Mutations
Mutation Rates and Outcomes
The mutation rates of DNA viruses estimate those of eukaryotic cells, yielding in theory one mutant virus in several hundred to many thousand genome copies. RNA viruses have much higher mutation rates, mayhap one mutation per virus genome re-create. Mutations can exist deleterious, neutral, or occasionally favorable. Simply mutations that practise not interfere with essential virus functions can persist in a virus population.
Phenotypic Variation by Mutations
Mutations can produce viruses with new antigenic determinants. The appearance of an antigenically novel virus through mutation is called antigenic migrate. Antigenically altered viruses may be able to cause disease in previously resistant or immune hosts.
Vaccine Strains from Mutations
Mutations can produce viruses with a reduced pathogenicity, altered host range, or altered target prison cell specificity only with intact antigenicity. Such viruses can sometimes be used equally vaccine strains.
Recombination
Recombination involves the commutation of genetic material between two related viruses during coinfection of a host cell.
Recombination by Independent Assortment
Recombination past independent assortment can occur among viruses with segmented genomes. Genes that reside on different pieces of nucleic acid are randomly assorted. This tin effect in the generation of viruses with new antigenic determinants and new host ranges. Development of viruses with new antigenic determinants through independent assortment is called antigenic shift.
Recombination of Incompletely Linked Genes
Genes that reside on the same slice of nucleic acid may undergo recombination. The closer ii genes are together, the rarer is recombination between them (partial linkage).
Phenotypic Variation from Recombination
Development of viruses with new antigenic determinants by either type of recombination may allow viruses to infect and cause affliction in previously immune hosts.
Vaccines through Recombination
Vaccine strains of viruses can be used to create recombinant viruses that deport extra genes coding for a specific immunogen. During viral vaccination, the replicating virus volition express the specific immunogen. Specific antibiotic production will be stimulated, and the host will be protected from the immunogen besides as from the vaccine virus.
Introduction
Viruses are simple entities, lacking an free energy-generating system and having very limited biosynthetic capabilities. The smallest viruses have merely a few genes; the largest viruses have as many as 200. Genetically, however, viruses have many features in common with cells. Viruses are subject field to mutations, the genomes of unlike viruses can recombine to form novel progeny, the expression of the viral genome can exist regulated, and viral cistron products tin collaborate. By studying viruses, nosotros can learn more than about the mechanisms past which viruses and their host cells function.
Genetic Modify in Viruses
This chapter covers the mechanisms past which genetic changes occur in viruses. 2 primary mechanisms are involved: mutation and recombination. Alterations in the genetic material of a virus may atomic number 82 to changes in the function of viral proteins. Such changes may issue in the creation of new viral serotypes or viruses of altered virulence.
Mutations
Mutations arise past one of iii mechanisms: (one) by the effects of concrete mutagens (UV calorie-free, x-rays) on nucleic acids; (two) by the natural behavior of the bases that make upward nucleic acids (resonance from keto to enol and from amino to imino forms), and (3) through the fallibility of the enzymes that replicate the nucleic acids. The first 2 mechanisms act similarly in all viruses; hence, the furnishings of physical mutagens and the natural behavior of nucleotides are relatively constant. However, viruses differ markedly in their mutation rates, which is due primarily to differences in the allegiance with which their enzymes replicate their nucleic acids. Viruses with high-fidelity transcriptases have relatively depression mutation rates and vice versa.
Mutation Rates and Outcomes
Deoxyribonucleic acid viruses have mutation rates similar to those of eukaryotic cells because, similar eukaryotic Dna polymerases, their replicatory enzymes have proofreading functions. The error rate for DNA viruses has been calculated to be x-eight to 10-11 errors per incorporated nucleotide. With this depression mutation rate, replication of even the virtually circuitous Deoxyribonucleic acid viruses, which have 2 × 10five to 3 × 105 nucleotide pairs per genome, will generate mutants rather rarely, mayhap once in several hundred to many thousand genome copies. The RNA viruses, however, lack a proofreading function in their replicatory enzymes, and some have mutation rates that are many orders of magnitude higher—10-iii to 10-4 errors per incorporated nucleotide. Even the simplest RNA viruses, which accept nigh vii,400 nucleotides per genome, volition generate mutants frequently, perchance as often every bit once per genome copy.
Not all mutations that occur persist in the virus population. Mutations that interfere with the essential functions of attachment, penetration, uncoating, replication, assembly, and release do not let misreplication and are rapidly lost from the population. All the same, because of the redundancy of the genetic code, many mutations are neutral, resulting either in no modify in the viral protein or in replacement of an amino acid by a functionally similar amino acrid. Only mutations that do not cripple essential viral functions can persist or become fixed in a virus population.
Phenotypic Variation by Mutations
Mutations that alter the viral phenotype just are not deleterious may be important. For case, mutation can create novel antigenic determinants. A mutation in the hemagglutinin gene of influenza A virus tin can give rising to a hemagglutinin molecule with an altered antigenic site (epitope) (Fig. 43-1). Provided the zipper function of the new hemagglutinin is intact, the mutant virus may be able to initiate an infection in an private immune to viruses expressing the previous hemagglutinin. For instance, from 1968 to 1979, mutations contradistinct 10 percentage of the amino acids in the influenza virus hemagglutinin serotype H3 molecule. This relatively pocket-size mechanism of antigenic modify through mutation, chosen antigenic drift, may allow a virus to outflank host defenses and cause disease in previously immune individuals.

Effigy 43-1
Mutation causing phenotypic (antigenic) variation. . Mutation of the codon for the hydrophilic amino acid serine to the codon for the hydrophobic amino acid phenylalanine tin change an epitope on the viral hemagglutinin protein and thereby alter its recognition (more than...)
Vaccine Strains from Mutations
Mutation has been a principal tool of virologists in developing attenuated live virus vaccines (Table 43-i). For instance, the Sabin vaccine strains of poliovirus were developed by growing polioviruses in monkey kidney cells. Mutation and choice produced variant polioviruses that were adapted for efficient replication in these cells. Some of the mutations in these variants affected the genes coding for the poliovirus coat proteins in such a way every bit to produce mutants unable to attach to homo neural cells but still able to infect human intestinal cells. Infection of human intestinal cells does not produce paralytic illness only does induce immunity. Poliovirus vaccine strains 1 and 2 have multiple mutations in the coat proteins and are very stable. The type iii vaccine strain is less stable and is field of study to back-mutations (reversions) that restore neural virulence. This vaccine strain therefore causes paralytic disease in ane out of every several meg vaccinated individuals. Despite the possibility of back-mutations, the generation and selection of adulterate viral mutants remains an important machinery for producing viral vaccines.
Recombination
Viral recombination occurs when viruses of two different parent strains coinfect the same host cell and collaborate during replication to generate virus progeny that take some genes from both parents. Recombination by and large occurs between members of the same virus type (e.1000., between two influenza viruses or between two herpes simplex viruses). Ii mechanisms of recombination have been observed for viruses: contained assortment and incomplete linkage. Either machinery can produce new viral serotypes or viruses with altered virulence.
Recombination by Contained Assortment
Contained assortment occurs when viruses that accept multipartite (segmented) genomes trade segments during replication (Fig. 43-ii). These genes are unlinked and assort at random. Recombination past independent assortment has been reported, for instance, for the influenza viruses and other orthomyxoviruses (viii segments of unmarried-stranded RNA) and for the reoviruses (10 segments of double-stranded RNA). The frequency of recombination by independent assortment is half dozen to twenty pct for orthomyxoviruses. Independent assortment between an animal and a human strain of influenza virus (come across Ch. 58) during a mixed infection can yield an antigenically novel influenza virus strain capable of infecting humans but carrying animal-strain hemagglutinin and/or neuraminidase surface molecules. This recombinant can infect individuals that are immune to the parent human virus. This mechanism results in an immediate, major antigenic modify and is called antigenic shift. Antigenic shifts in flu virus antigens tin give ascension to pandemics (worldwide epidemics) of influenza. Such antigenic shifts have occurred relatively frequently during recent history (Tabular array 43-ii). Because the number of different serotypes of hemagglutinin and neuraminidase are limited, a given strain reappears from time to time. For example, the H1N1 flu virus strain was responsible for the 1918 to 1919 influenza pandemic that acquired xx million deaths. The same virus also caused pandemics in 1934 and in 1947, so disappeared later on 1958 and reappeared in 1977. The reappearance of virus strains subsequently an absence is believed to be the outcome of recombinational events involving the independent assortment of genes from two variant viruses.

Effigy 43-2
Recombination by contained assortment during dual infection. Later infection of a cell with two viruses with 2 or more genetic segments ("chromosomes"), reassortment of the replicated segments can occur. Independent assortment results (more than...)
Table 43-2
Antigenic Shifts Resulting from Reassortment of Genome Segments.
Recombination of Incompletely Linked Genes
Recombination too occurs between genes residing on the aforementioned piece of nucleic acid (Fig. 43-3). Genes that more often than not segregate together are called linked genes. If recombination occurs between them, the linkage is said to exist incomplete. Recombination of incompletely linked genes occurs in all DNA viruses that have been studied and in several RNA viruses.
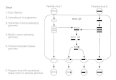
Figure 43-iii
Recombination by interruption-rejoin of incompletely linked genes. . The genetic interaction of DNA viruses tin result in break-rejoin recombination, in which the two DNA molecules of different viruses break and and so cross over. Break-rejoin recombination results (more...)
In DNA viruses, as in prokaryotic and eukaryotic cells, recombination betwixt incompletely linked genes occurs past means of a break-rejoin mechanism. This mechanism involves the actual severing of the covalent bonds linking the bases of each of the two Deoxyribonucleic acid strands in a Deoxyribonucleic acid molecule (Fig. 43-3). The severed Deoxyribonucleic acid strands are then rejoined to the DNA strands of a dissimilar DNA molecule that has been broken in a like site. Recombination rates for herpesviruses, which are DNA viruses that replicate in the nucleus of infected cells, estimate those expected for a eukaryotic genome of the size of the herpesvirus genome. Herpesviruses have an average recombination frequency of x to 20 per centum for whatever 2 loci. Nonetheless, the rate of recombination betwixt a specific pair of genetic loci depends on the distance between them and varies from less than 1 percent to approximately 50 percent. Measurement of the recombination frequencies for different loci can exist used to map the virus genome. In this blazon of genetic map, loci with high recombination frequencies are far apart and loci with depression recombination frequencies are close together.
Recombination has been shown to occur in several positive-sense single-stranded RNA virus groups: retroviruses, picornaviruses, and coronaviruses. That is initially surprising, every bit recombination betwixt RNA molecules has not been observed in prokaryotic or eukaryotic cells. In retroviruses, recombination really occurs at the point in replication when the retrovirus genome is in a Dna form and takes place by the same break-rejoin machinery as in cells and DNA viruses. Recombination can occur both between two related retroviruses and between the retrovirus Deoxyribonucleic acid and the host cell DNA. Recombination between two retroviruses gives rise to novel viral progeny with reassorted genes. Recombination betwixt retroviruses and the host jail cell can give rise to novel viral progeny that carry nonviral genes. If these host genes code for growth factors, growth factor receptors, or a number of other specific cellular proteins, the recombinant retroviruses may be oncogenic (run into Ch. 47).
In picornaviruses and coronaviruses, recombination takes identify at the level of the interaction of the viral RNA genomes and is not believed to occur by a break-rejoin mechanism. The mechanism is currently believed to be a re-create-choice mechanism (Fig. 43-4). Re-create-choice may occur in these RNA viruses because the viral RNA polymerase binds to only a few bases of the template RNA at any ane fourth dimension. Such a weak interaction of the polymerase with the template RNA would permit the polymerase, carrying its RNA strand, to disassociate from the original template nucleic acid strand and then associate with a new template RNA strand. Recombination frequencies in the range of 0.2 to 0.4 pct have been reported. Therefore, the efficiency of this mechanism of recombination is depression.
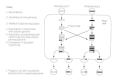
Effigy 43-iv
Recombination by copy-choice of incompletely linked genes. The genetic interaction of certain RNA viruses can result in copy-option recombination. In this machinery, the polymerase begins replicating RNA template. Past an unknown mechanism, which may involve (more...)
Phenotypic Variation from Recombination
As mentioned above, viral recombination is important considering information technology tin can generate novel progeny viruses that express new antigenic and/or virulence characteristics. For example, the novel progeny viruses may have new surface proteins that let them to infect previously resistant individuals; they may have altered virulence characteristics; they may take novel combinations of proteins that make them infective to new cells in the original host or to new hosts; or they may comport material of cellular origin that gives them oncogenic potential.
Vaccines and Cistron Therapy through Recombination
Recombination is being used experimentally past virologists to create new vaccines. Vaccinia virus, a DNA virus of the poxvirus grouping, was used as a live vaccine in the eradication of smallpox. Recombinant vaccinia viruses are being developed that carry vaccinia virus Deoxyribonucleic acid recombined with DNA from other sources (exogenous DNA) (Fig. 43-5). For example, vaccinia virus strains carrying Dna coding for bacterial and viral antigens accept been produced. It is expected that subsequently vaccination with the recombinant vaccinia virus, the bacterial or viral antigen (immunogen) will be produced. The presence of this immunogen will then stimulate specific antibody production past the host, resulting in protection of the host from the immunogen. Studies with these live, recombinant vaccinia viruses are currently under mode to decide whether inoculation of the skin with the recombinant virus can induce a protective host antibiotic response to the bacterial or viral antigens. Other studies are investigating the use of live, recombinant adenoviruses containing bacterial or viral genes to infect the gastrointestinal tract and induce both mucosal and systemic amnesty.
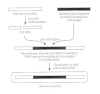
Effigy 43-5
Evolution of recombinant vaccinia virus for immunization against cholera toxin. Vaccinia virus genomic Deoxyribonucleic acid is cut with an endonuclease. A specific sequence of DNA (with appropriate regulatory sequences) coding for a protein (due east.one thousand., cholera toxin) to (more...)
In a like fashion, recombinant viruses are besides beingness developed that carry normal human genes. It is envisioned that such recombinant viruses could exist useful for factor therapy. Target diseases for gene therapy span a wide range, including diabetes, cystic fibrosis, severe combined immunodeficiency syndrome, etc. Indeed, treatment of cystic fibrosis patients with replication deficient, recombinant adenoviruses begetting a normal copy of the cystic fibrosis transmembrane regulator gene has already been approved.
If these studies give positive results, such directed generation of recombinant viruses may get an important tool in the development of vaccines and for gene therapy.
References
-
Cox NJ, Brammer TL, Regnery HL. Influenza: global surveillance for epidemic and pandemic variants. Eur J Epidemiol. 1994;ten:467. [PubMed: 7843358]
-
Gao L, Chain B, Sinclair C. et al. Immune response to human papillomavirus type 16 E6 factor in a live vaccinia vector. J gen Virol. 1994;75:157. [PubMed: 7509369]
-
Kingdom of the netherlands J, Spindler K, Horodyski F. et al. Rapid development of RNA genomes. Science. 1982;215:1577. [PubMed: 7041255]
-
Honess RW, Buchan A, Halliburton IW, Watson DH. Recombination and linkage between structural and regulatory genes of herpes simplex virus type I: study of the functional organization of the genome. J Virol. 1980;34:716. [PMC free article: PMC288761] [PubMed: 6247508]
-
Palese P, Young JF. Variation of influenza A, B, and C viruses. Science. 1982;215:1468. [PubMed: 7038875]
-
Paoletti E, Perkus ME, Piccini A. Live recombinant vaccines using genetically engineered vaccinia virus. Antiviral Res, suppl. 1985;1:301. [PubMed: 3866514]
-
Radding CM. Homologous pairing and strand exchange in genetic recombination. Annu Rev Genet. 1982;16:405. [PubMed: 6297377]
-
Romanova LI, Blinov VM, Tolskaya EA. et al. The primary structure of crossover regions of intertypic poliovirus recombinants: a model of recombination between RNA genomes. Virology. 1986;155:202. [PubMed: 3022471]
-
Schaffer PA, Tevethia MJ, Benyesh-Melnick M. Recombination between temperature sensitive mutants of herpes simplex virus type 1. Virology. 1974;58:219. [PubMed: 4362547]
-
Scholtissek C. Source for influenza pandemics. Eur J Epidemiol. 1994;10:455. [PubMed: 7843354]
-
Siegfried W. Perspectives in gene therapy with recombinant adenoviruses. Exp Clin Endocrinol. 1993;104:7. [PubMed: 8477824]
-
Smith FI, Palese P: Variation in flu virus genes: epidemiological, pathogenic, and evolutionary consequences. p. 319. In Krug RM (ed): The Influenza Viruses. Plenum, New York, 1989 .
Source: https://www.ncbi.nlm.nih.gov/books/NBK8439/#:~:text=They%20undergo%20subtle%20genetic%20changes,information%2C%20creating%20a%20novel%20virus.
Posted by: holguinexquours.blogspot.com
0 Response to "How Can Viruses Change The Genetic Makeup"
Post a Comment